
Scientific journal
European Journal of Natural History
ISSN 2073-4972
ИФ РИНЦ = 0.204
DEGRADATION OF PHOTOSYNTHETIC PIGMENTS AND CHANGES IN THE CONTENT OF FREE PROLINE AND GUAIACOL PEROXIDASE UNDER CONDITIONS OF DROUGHT AND SALINIZATION

Soybean is an important agricultural crop, rich in proteins, fats, micro-, and macro-elements. The demand for soybean crop is growing all over the world. Despite the development of high-yielding varieties and increased productivity, the loss is not small. Most of the losses, which account for about 1/5 of the total product, are due to abiotic stress factors [1].
According to the data obtained, soy is a plant that is more susceptible to the influence of abiotic stress factors, especially drought [2]. The influence of stress factors that create a lack of water is more often manifested in the growth and development phase of ontogenesis. Thus, as a result of water shortage, the leaf surface of plants shrinks, the height decreases, the absorption of nutrients decreases, the amount of beans decreases.
Therefore, the study of the physiological and biochemical aspects of the effects of drought and salinity, which are characteristic of the Azerbaijani climate, is a priority. The development of the early diagnostics based on the physiological and biochemical indices of stress tolerance is of great importance in the selection of tolerant varieties and forms.
Materials and methods of research
Five genotypes of soybean (Bravo, Beyson, Agroyol, Regale, Sinara) cultivated in the experimental field of the Institute of Genetic Resources were used in the research. Tolerance to drought and salinity stresses was evaluated based on changes in the chlorophyll content, free proline content and the activity of guaiacol peroxidase in soybean leaf samples. For this purpose, top leaf samples taken from field experimental variants of soybean genotypes were exposed to salinity and drought stresses in the laboratory. Saline and arid environments were created using 1.5% NaCl (11 atm) and 8.7% (7 atm) sucrose solutions following the stress tolerance limit [3]. The level of chlorophyll (a + b) in leaves was measured using a portable device SPAD-502Plus. The device determined the spectral absorption in two ranges, and based on the data obtained, calculated the indexed value of the chlorophyll content in the leaves.
The proline content in leaves of stress-exposed leaves was determined using the Bates method [4]. The optical density of proline was measured at 520 nm using a spectrophotometer (UV-3100 PC).
The definition of proline was based on known methodology. The 500 mg wet plant biomass was squashed with 20 ml of 3% sulfosalic acid and filtered the thick blue filter until it was homogeneous. After pouring of 2ml from filtrate to test glass, also 2ml reagent (1,25g ninhydrin, 30 ml of vinegar acid and 20ml 6M phosphoric acid mixture), 2 ml of vinegar acid was added and placed in boiling water bath for 1 hour after thorough mixing. After that, the test glasses were taken out and cooled to + 4 ° C. 4 mL toluene was added to the reaction mixture and rested after shaking it for 15 seconds. The colored solution is gathered on top of the toluene and its color may vary from light pink to dark red, depending on the amount of proline. The top coat was taken up with a pipette and poured into the cuvette and measured the optical density of toluene.
Determination of peroxidase activity by spectrophotometry is based on measuring the optical density of products formed during the guaiacol oxidation reaction. After crushing the leaves (200 mg) in a porcelain cup with a small amount (5-10 ml) of phosphate buffer (pH 5.4), the mixture was centrifuged for 10 minutes at a speed of 4000-5000 rpm. The optical density of the reaction mixture consisting of 0.5 ml H2O2, 0.5 ml substrate (guaiacol), 1.5 ml phosphate buffer, 0.5 ml supernatant (enzymatic plant material) was measured on a spectrophotometer (UV-3100 PC) for 1 min, at a wavelength of 470 nm [5].
Results of the research and discussions
In order to assess the condition and working capacity of pigment systems in green plants under drought and salinity conditions, it is important to determine the chlorophyll content of photosynthetic pigments.
The level of chlorophyll (a+b) in the leaves was measured using a portable device SPAD-502Plus. The device determined the spectral absorption in two ranges, and based on the data obtained, calculated the indexed value of the chlorophyll content in the leaves.
According to the SPAD method, chlorophyll (a+b) ranged between 27.2 and 33.5 in the control plants. The maximum pigment content was found in the Bravo specimen, while the minimum value was observed in the stressed plants of the Beyson variety.
Under drought conditions, the total amount of photosynthetic pigments varied in the range of 25.7-32.4, the maximum value was observed in the Regale specimen, and the minimum value was detected in the Beyson variety. Chl (a+b) was found to decrease by 5.6 (17%) and 1.5 (5.5%) under drought in the Bravo and Beyson varieties, respectively. Whereas, a slight decrease of this parameter – 0.6 was detected in the Agroyol specimen. In the Regale specimen, chl (a+b) increased by 4 (14%), whereas, in the Sinara variety only a slight increase occurred.
Under salinity, the total amount of photosynthetic pigments changed in a wider range of 24-33,1. Thus, the highest value of chlorophyll was obtained in the Regale variety, and the lowest value was found in the Beyson variety. Compared to the control value, chlorophyll (a + b) amounted to 116.5% (Regale), 109.4% (Agroyol), 90% (Sinara), 88.2% (Beyson) and 81.5% (Bravo) under salinity.
The visual physiological observation of soybean specimens 1 day after their exposure to drought stress corresponding to the tolerance limit showed that the stressed variants of Bravo, Beyson, and especially Agroyol maintained turgor, and the leaves did not turn yellow or sag. Although a slight turgor loss was observed in the Regale specimen, the effect of stress was more pronounced in the Sinara specimen. According to the physiological observations, Sinara did not tolerate drought lasted 48 hours. In contrast to drought stress, which sharply slowed growth and development, salinity had milder effects on the specimens. Thus, even the 48-hour effect of salinity did not lead to the death of any variety.
The proline content in control plants varied in the range of 0.31-1.36 µM/g (Figure 2).
According to the upper limit of tolerance, the free proline content in soybean specimens exposed to drought stress for 24 hours varied in the range of 1.96–9.0 µM/g. The obtained values increased by 5.66 times (Sinara) – 10.26 times (Agroyol) compared to control.
The proline content changed in the range of 1.51-3.18 µM/g and this parameter increased 2.21 times (Bravo) – 4.9 times (Beyson and Aqroyol) compared to control after diurnal exposure to salinity stress.
The Sinara variety-sample from soybean experiment options did not withstand drought stress for 48 hours. The amount of free proline in the remaining samples increased slightly, amounting to 1,96-11,31 µM /g. This increase was 6.32 (Beyson) – 17.96 (Agroyol) times for control, compared to 1-day drought-1.0 (Beyson) – 1.75 (Agroyol) times (Figure 3).
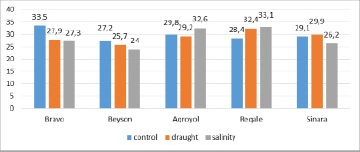
Fig. 1. The amount of chlorophyll (a+b) in soybean samples
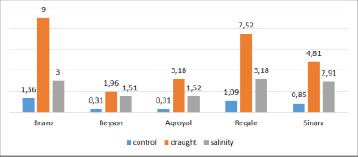
Fig. 2. The amount of free proline in the leaves (µM /g) after 24 hours of stress on soybeans plants
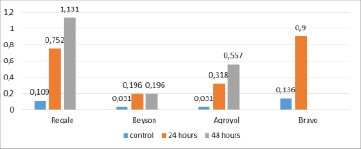
Fig. 3. The amount of free proline in the leaves after drought stress, which affects 24 and 48 hours in soybean plants
In plants exposed to various abiotic stress factors, an increase in proline occurs as a general physiological response. The increase in the proline content can be attributed to its de novo synthesis, reduced degradation, low utilization. While the polyfunctionality of proline is still the subject of debate, this amino acid is believed to regulate the oxidation-reduction potential of cells, participates in the capture of free radicals, and, first of all, performs an osmoprotective function by regulating cell water balance. Besides, it forms chelate compounds with heavy metals and even induces the expression of genes of antioxidant enzymes [6]. Probably, proline acts as a signal element associated with stress and is involved in the formation of tolerance to some stress factors.
Proline accumulation may be activated at a certain stage of the response to stress and a certain stress degree. The accumulation of proline under mild stress can be considered as an adaptive response in sensitive plants, while the accumulation of large amounts of proline was recorded in tolerant plants under severe stress [7].
Proline is known to act as an osmoprotectant in the plant adaptation to drought and salinity. Accumulation of proline in large quantities leads to an increase in intracellular osmolarity, thereby maintaining water balance and important turgor under water deficiency [8].
Guaiacol-peroxidase (PO). There was a decrease in guaiacol-peroxidase activity in all soybean experiment variants after 24 hours of drought. Thus, the activity of PO was in the range of 0.106 – 0.224 and decreased by 1.04 (Beyson) – 1.83 times (Bravo) (Figure 4).
Determination of enzyme activity after 24- and 48-hour exposure to salt stress on the leaves of soybean samples showed that the activity of guaiacol peroxidase decreased during short-term exposure, but increased sharply after prolonged exposure. in plants aged in a salt medium for 1 day, the activity of guaiacol peroxidase varied in the range of 0.095-0.284 and was 0.43 (Agroyol) – 1.45 (Sinara) parts from the control. After 2 days of salinization, the enzyme became more active and received values in the range of 0.376-1.006 and increased by 1.62 times (Beison) – 5.15 times (Bravo) in relation to control, 2.41 times (Sinara) – 5.59 times (Bravo) in relation to the first day of stress (Figure 5).
To maintain viability in unfavorable conditions, plants created effective antioxidant mechanisms with two shoulders: enzymatic components from the family of peroxidase, catalase and SOD, and non-enzymatic components, consisting of ascorbic acid, α-tocopherol, carotenoids, flavonoids, proline-osmolite. These two components jointly perform such an important function as neutralizing the active forms of oxygen, which are absorbed in considerable amounts during stress and can pose serious dangers for the normal survival of the plant organism [9].
Determination of free proline content and guaiacol peroxidase activity in soybean leaves after 24 hours of exposure to drought stress showed that, although the content of proline amino acids increased in all variants, the enzyme activity decreased. The same pattern was observed with 24-hour exposure to salt stress (Figure 6).
According to literature data [9, 10] enzymatic and non-enzymatic arms of the antioxidant protection system work mutually to establish tolerance to unfavorable environment in plants and to mitigate the pest effects of radical forms of oxygen. The results we have received in this regard are consistent with the literature data and support the ideas addressed above.
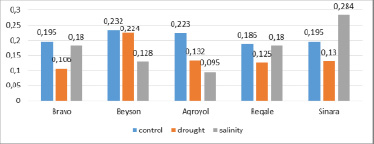
Fig. 4. Activity of guaiacol-peroxidase in leaves after 24 hours of stress on soybean plants
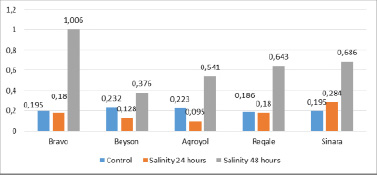
Fig. 5. Determination of guaiacol-peroxidase activity in leaves after 24 and 48 hours of salinity stress in soybean plants
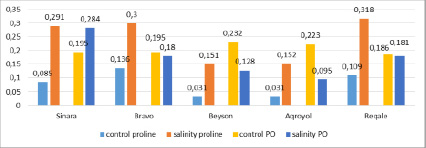
Fig. 6. The amount of proline and the activity of guaiacol-peroxidase (PO) in the leaves of soybean plant during 24-hour exposure of salinity
Recently, more attention has been added to the antioxidant function of proline. Its structure allows to consider the inactivation of radical forms of oxygen in a certain way. As can be seen from the results of the research works, proline plays an important role in the inactivation of hydroxyl radical and is also involved in the detoxification of hydrogen peroxide when the amount of proline in the cell is more [11, 12]. Since enzymatic antioxidants can neutralize hydrogen peroxide, superoxide radicals, singlet oxygen, but cannot neutralize the hydroxyl radical, and also because of the high content of proline in cell complexes, such a mechanism is a convenient way to protect against free radicals. On the other hand, information is provided about the location of a large amount of free proline in the cytoplasm and its protection of antioxidant enzymes from various damages and possible consequences of denaturation [13].
It is known that proline acts as an osmoregulator in the adaptation of plants to drought and salinization. Its accumulation in large quantities in the cell leads to an increase in intracellular osmolarity and, thus, ensuring water balance, water potential and, importantly, turgor in conditions of water scarcity [8].
Conclusions
Referring to the results obtained, it can be noted that the effect of dehydration and salinization stress factors to soybean in experimental variants led to degradation of photosynthetic pigments. Given the greater accumulation of free proline from strong stress effects in resistant plants, we can conclude that the Beyson and Agroyol samples are relatively resistant to stress factors.
Библиографическая ссылка
Ибрагимова З.Ш., Абдуллаева Л.С., Алиев Р.Т. DEGRADATION OF PHOTOSYNTHETIC PIGMENTS AND CHANGES IN THE CONTENT OF FREE PROLINE AND GUAIACOL PEROXIDASE UNDER CONDITIONS OF DROUGHT AND SALINIZATION // European Journal of Natural History. 2022. № 3. ;URL: https://world-science.ru/en/article/view?id=34270 (дата обращения: 28.07.2026).