
Scientific journal
European Journal of Natural History
ISSN 2073-4972
ИФ РИНЦ = 0.204
EVALUATION OF PHYSIOLOGICAL AND BIOCHEMICAL PARAMETERS OF BREAD WHEAT (T. AESTIVUM L.) SAMPLES UNDER STRESS

Since ancient times Azerbaijan has been the homeland for many types of cereals, including some species of wheat. The natural condition of our republic is favorable for the cultivation of grain crops. However, seasonal droughts, high temperatures, salinity and natural moisture instability are negative factors that impede high productivity. Therefore, due to climate changes, developing tolerant varieties and forms of agricultural plants, especially cereals, is extremely actual. Determination and improvement of plant tolerance to high temperature and low humidity conditions are to be based on deep knowledge of physiological and biochemical properties of plants. The comparative study of physiological and biochemical properties of wheat samples belonging to various species contributes to the establishment of their tolerance limits to abiotic stress factors, assessment of adaptation potential, thereby resolving the problem of the regulation of the productivity processes in cereals.
The effect of stress factors, such as drought, salinity, high temperature, first of all, causes water deficiency and the formation of free radicals, wherein strong defense mechanisms are launched to regulate cell water supply. One of these mechanisms is the accumulation of low molecular weight compounds of the antioxidant defense system, especially, proline. Proline effectively triggers adaptation mechanisms under stress conditions and beeing a multifunctional stress metabolite, according to modern concepts, besides osmoprotective functions, it performs also chaperone, antioxidant, signal-regulator and other functions (1, 2).
On the other hand, seeds that better tolerate the adverse conditions of accelerated aging, are known to be more tolerant to abiotic stressors during germination. There is information on the relationship between aging with impaired physiological and biochemical processes (3, 4, 5) and the accumulation of growth inhibitors and toxic metabolic products. At the heart of the damage that leads to the aging of seeds and reduce their stress tolerance is the generation of free radicals, which initiates lipid peroxidation (6). These reactions do not require high moisture content and their products can accumulate in air-dry seeds, which leads to oxidative damage to the membranes, proteins and DNA. According to modern concepts, some amino acids, including proline, initiate the activation of adaptive mechanisms (7).
The aim of research
The aim of this study was to determine the tolerance of wheat samples of various species to abiotic stress factors (drought, salinity and artificial aging), based on their physiological and biochemical parameters.
Materials and research methods
The objects of the study were seeds of 9 samples (7 varieties) of bread wheat: K-3 T.aestivum var.milturum AL., K-6 T.aestivum var.miltrum, K-17 T.aestivum var.erythrospermum Al., K-24 T. aestivum var.ferrugineum AL, K-27 T.aestivum var.ferrugineum Al., K-35 T.aestivum var.erythroleucon Korn., K-43 T. aestivum var.barbarossa AL., K-65 T.aestivum var.leucospermum Korn., K-72 T. aestivum var.cianotrics Korn. Stress factors were accelerated seed aging, dehydration and salinization of seedlings. To imitate the duration of the storage of seeds, the method of artificial aging was used. This method involves a 3-day incubation of seeds at elevated relative humidity (95 %) and air temperature (40 °C) (3 Smolikova G.N, 2014), which allows simulating the effects of adverse factors and predicting their effect on the tolerance of seeds of various varieties and plant samples. Viability was evaluated according to the test of the seed germination ability under laboratory conditions, expressed in the percentages of the total number (n) G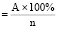 , where A – number of sprung seeds.
, where A – number of sprung seeds.
For biochemical analysis, plants were grown in the phytotron under the following conditions: 21 °-22 °C temperature, 60 % humidity, 16h/8h photoperiod, 10000 lux illumination. According to the tolerance limits (8), 5-day-old seedlings were exposed to drought and salt stresses (using 20 atm sucrose and 16 atm NaCl solutions) for 24 hours and artificial aging for 3 days. Then the proline amount was determined in leaves using the Bates method (9). Comparative analysis was performed with 6-day-old seedlings. Samples were taken at the same time of day and plants grown under normal conditions were used as control.
The definition of proline was based on known methodology. The 0.5g wet plant biomass was squashed with 20 ml of 3 % sulfosalic acid and filtered the thick blue filter until it was homogeneous. After pouring of 2ml from filtrate to test glass, also 2ml reagent (1,25g ninhydrin, 30 ml of vinegar acid and 20ml 6M phosphoric acid mixture), 2 ml of vinegar acid was added and placed in boiling water bath for 1 hour after thorough mixing. After that, the test glasses were taken out and cooled to + 4 °C. 4 mL toluene was added to the reaction mixture and rested after shaking it for 15 seconds. The colored solution is gathered on top of the toluene and its color may vary from light pink to dark red, depending on the amount of proline. The top coat was taken up with a pipette and poured into the cuvette and measured the optical density of toluene. The amount of proline is calculated by the following formula (10):
C = E•k•V/m
C-the amount of proline (μM/g), E-optical density, k- coefficient of calibration curve (217.49), V-volume of extract (ml), m-plant material weight (g).
Research results and discussion
To assess functional impairment of seed viability during accelerated aging, we used an integral index such as their germination (Fig. 1). Under optimal conditions for germination, the seed germination ability of 9 bread wheat varieties ranged from 84.0 % to 100.0 %. The study of the germination ability of 9 bread wheat varieties subjected to accelerated aging of seeds showed that 3-day aging reduced the germination of seeds from different samples to different degrees. Thus, sharp declines – 12 % and 46.5 % – in the germination ability was observed in the samples K-43 var.barbarossa AL. and K-72 – var. cainotrics Korn, respectively. Whereas, in the samples K-17 var.erythrospermum Al., K-24 – var. ferrugineum AL., K – 27 – var.ferrugineum AL. и K – 35 var.erythroleucon Korn. only 2.0 % decline was detected, which confirms higher tolerance of them to accelerated aging.
Accumulation of proline is considered to be the first response induced by stress in plant organisms. In some investigations, the proline accumulation ability of cells was used as a selective trait in the evaluation of drought and salt tolerance of species and varieties (11).
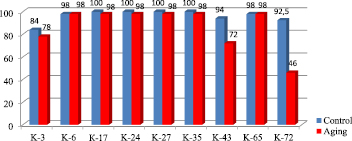
Fig. 1. Germination ability of the accelerated aging seed bread wheat varieties (K-3 T.aestivum var.milturum AL., K-6 T.aestivum var.miltrum, K-17 T.aestivum var.erythrospermum Al., K-24 T. aestivum var.ferrugineum AL, K-27 T.aestivum var.ferrugineum Al., K-35 T.aestivum var.erythroleucon Korn., K-43 T. aestivum var.barbarossa AL., K-65 T.aestivum var.leucospermum Korn., K-72 T. aestivum var.cianotrics Korn..)
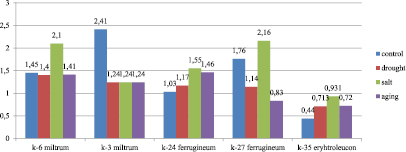
Fig. 2. The proline accumulation abilities of bread wheat varieties samples (K-6 T.aestivum miltrum, K-3 T.aestivum var.milturum AL., K-24 T. aestivum var.ferrugineum AL, K-27 T.aestivum var.ferrugineum Al., K-35 T.aestivum var.erythroleucon Korn.) exposed to stress factors (µM/mg)
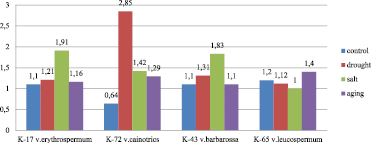
Fig. 3. The proline accumulation abilities of bread wheat varieties samples (K-17 T.aestivum var.erythrospermum Al., K-72 T. aestivum var.cianotrics Korn., K-43 T. aestivum var.barbarossa AL., K-65 T.aestivum var.leucospermum Korn.) exposed to stress factors (µM/mg)
We observed genetic variations in the proline accumulation abilities of samples exposed to stress factors. The changes in the proline amount under stress were studied in 9 samples (Fig. 2, 3). Wide variations were observed in all variants. In control variants proline amount ranged from 0.44 µM/mg to 2.24 µM/mg. Minimum and maximum limits of this parameter were found in the samples K-35 var. erythroleucon and, K-3 var. miltrum, respectively.
The use of resource substances is the first response of plant organisms to stress effects. Resource substances involve special enzymes, including antioxidant enzymes and low-molecular metabolites (for example, proline). Then the synthesis of the components required for the activation of the defense system intensifies. There are many reports on this subject (12). Some of our results are related to the issue and consistent with the literature information. Thus, in plants of the var. miltrum K-3 samples, the proline amount decreased 2 times under all 3 stress factors which is 51.5 % of the control value. This means that during one day of stress, 1.17μM proline was consumed per 1 mg of fresh weight and this value is the largest one among other samples manifesting a decreased proline amount. It is suggested that this time is less than required for the synthesis of low-molecular compounds of the antioxidant system, in our case proline. Similar results were obtained for the seedlings of var. ferrugineum K-27 and var. leucospermum K-65. Thus, in the first variant, the proline amount was 64,8 % of the control value under drought and 47.2 % under artificial aging. In the second variant 93 % under drought and 83 % under salinity.
The proline amount ranged from 0.71 µM/mg to 2.85 µM/mg under drought. Minimum and maximum values were found for var. erythroleucon K-35 and var. cainotrics K-72, respectively. The proline amount increased 4.45 times compared with control and became maximum in the var. cainotrics K-72 plants. The samples var. miltrum K-6, var. erythrospermum K-17, var. ferrugineum K-24 showed the closest to control values for the proline amount under drought.
The proline amount changed between 0.93 µM/mg and 2.16µM/mg under salinity. The smallest value was detected in var. erythroleucon K-35. In var.erythroleucon K-35 and var. cainotrics K-72 the proline amount was 2 times higher compared with control. In the seedlings of var. leucospermum K-65 the proline amount was 17 % less (83.3 % of the control value) compared with control, which is attributed to 0.2 µM/mg consumption of proline reserves and a delay of the synthesis of new proline.
In plants exposed to artificially aging treatment, the proline amount was in the range of 0.72 to 1.46 µM/mg. The smallest value was observed for var. erythroleucon K-35 and the largest value for var. ferrugineum K-24. Thus, these values increased 1.64 and 1.42 times, respectively, compared with control. The value closest to control was registered for the var. barbarossa K-43 samples.
It is noteworthy that multiple increases in the proline amount under drought, salinity and accelerated aging occurred in samples, which control variants had low levels of the constitutive proline amounts (var.erythroleucon K-35, var. cainotrics K-72). Probably, in order to tolerate stress, more active functioning of the defense system is required in these plants. Proline was found to accumulate to a greater extent (as a result of its synthesis, as well as degradation of proteins (13) in sensitive plants. In stress exposed seedlings of var. cainotrics K-72, the proline amount was 4.45 times higher compared with control. In contrast, the proline amount in the tolerant plants was almost as in control. This can be attributed to higher water retention and reparation abilities of tolerant plants (14). In tissues of plants sensitive to adverse environmental conditions, the proline amount increases faster and becomes more compared with tolerant plants. However, a sharp difference between proline amounts in stress exposed and control variants of tolerant plants can be detected only under severe stress conditions (15).
We observed different responses to stress factors of the defense system in our researches. Our results suggest that under stress factors adaptation mechanisms are needed to function more effectively in sensitive plants having fewer proline reserves. In some samples, stress caused the expenditure of the proline reserves and activation of the antioxidant defense system (var. miltrum K-3, var. ferrugineum K-27, var. leucospermum K-65). Other samples showed tolerance to a various extent under drought, salinity and artificial aging.
Conclusion
A comparative assessment of the resistance of the studied wheat samples seeds to aging and the response of plants to the effects of drought and salinity showed that, in terms of seed germination after accelerated aging and the content of proline in the leaves of plants, the К-6 var. miltrum, K-17 var.erythrospermum, К-24 var. ferrugineum wheat showed greater stability compared to the other studied samples. The least stable was the K-72 T. aestivum L. var.cainotrics Korn. sample.
Библиографическая ссылка
Ibrahimova Z.Sh., Mammadova S.A., Hasanova G.I., Aliyev R.T. EVALUATION OF PHYSIOLOGICAL AND BIOCHEMICAL PARAMETERS OF BREAD WHEAT (T. AESTIVUM L.) SAMPLES UNDER STRESS // European Journal of Natural History. 2020. № 1. ;URL: https://world-science.ru/en/article/view?id=34030 (дата обращения: 11.06.2026).