
Scientific journal
European Journal of Natural History
ISSN 2073-4972
ИФ РИНЦ = 0.204
BIOCENOSIS, BIOCENOTIC DIVISION OF MAMMALS IN NATIONAL NATURAL PARK CHON-KEMIN AND CHARACTERISTIC OF CERTAIN MAMMALS OF THIS PARK

Recently one of the main directions in biology is research in the area of preserving bioviriety, genofond of species and their bio-efficiency, as well as their biocenotic division. Biovariety is studied at different levels of lively systems – species level, level of population and biocenosis.
History of studying biovariety of mammals in Kyrkyzstan began from classification and description of species in the second half of XIX century. The earliest data on mammals of Northern Tjan-Shan can be found in works by Pallas. Some data on mammals was collected in works by N.A. Severtsov (1857-1879), V.N. Shnitnikov (1936), N.V. Minin (1938). Later the research was resumed by S.I. Ognev (1928, 1947), A.T. Toktosunov (1958, 1972, 1984), A.I. Yanushevich, M. Aisin, A.K. Kydyraliyev (1972), E.D. Shukurov (1966, 1989), as well as other scientists who contributed greatly into research of spreading, morphology, nutrition, reproduction, and systematics of mammals of Northern Tjan-Shan. Due to inhabiting different biocenosis, populations of species of mammals adapted to the most various ecologic conditions. In this regard important is studying influence of different ecologic factors upon intraspecific variability of isolated populations. The importance of studying variability of populations in natural environment was specifically outlined by S.S. Schwarz (1980). As separate populations have their own biovariety due to variability under the influence of natural environment, these isolated populations serve as certain “natural laboratory” (A.V. Yablokov, 1987), and nature itself carries out experiments. As mammals are a part of lively nature, their comprehensive research and collection of scientific data continues.
Materials and research methods
5 species of mammals of Chon-Keminskiy population were studied: сommon pipistrelle, grey hamster, Tjan-Shan birch mouse, Tjan-Shan forest vole, narrow-skulled vole (10 specimen of each). Preparations of mytopic chromosomes were made from cells of bone marrow according to the methodic [20]. More than 2000 chromosome sets of each species were studied. Objects of field research for defining biocenosis of national natural park Chon-Kemin and ecologically-dominant types were expeditionary observations of mammals, carried out in the region of Chon-Keminskiy national natural park (1997-2018).
Research results and discussion
Dynamics and division
of mammals in biocenosis
For the first time biocenosis of national natural park Chon-Kemin were divided. While dividing territories of National natural park Chon-Kemin into biocenosis we must consider vertical zonality of mountatins where the corresponding local areas of vegetation are formed. According geobotanic map and data of K. Isakov (1959), as well as our data, territory of the park was divided into 4 biocenosis.
As valley Chon-Kemin is a mountain region, its vertical chains consist of different biotopes. In most cases a certain species cannot be found around the whole biotope, and, therefore, is spread in that local area of the biotope that corresponds to all ecologic requirements of this species. This area of the biotope was named by N.V. Minin (1938) as “space of life” and characterized by the fact that it has a limit of biological optimum for population of species. There are very few species of mammals that are satisfied with one life space, and in most cases they use the whole biotope. Certain eurytopic species of mammals (Mus musculus, Cricetulus migratorius, Ellobius talpinus, etc.) live in different ecological conditions, from by-mountain areas to mountain biotopes, for such species as Ochotona rutile stenotopity is typical, and they inhabit only biotopes of rocky deposits. “Space if life” for a species in certain cases can be spread over the whole realm or unevenly. In such conditions population of a species lives in similar biotopes, and sometimes they relocate from one biotope to another and create groups of specimen. While analyzing fauna composition of mammals by each biocenosis, this list does not describe in full composition of biocenosis. It seems that the main attention should be devoted to those populations of species of mammals for which high density in biocenosis is typical, so populations of these species form a “background” of this biocenosis. N.V. Minin (1938) underlines that ecologically-plastic species mostly live in one geographic realm in different biotopes, and they form heterogenous populations of species, in other words, ecotopes. Serafinski Wlodzimierz (1967) points out that many sub-species should be studied as ecologic populations, and sub-species – ecotopes are biomarkers of special characteristics of environment of the given realm, and these sub-species are geo-ecotopes. Supporting ideas of this scientist, we consider it necessary to study sub-species of mammals that inhabit Chon-Keminskiy national natural park as ecotopes of species, but not in status of sub-species. 37 species of ecotope mammals live at the territory of national natural park.
Biocenosis of low-mountain steppe
This biocenosis of low-mountain steppe is located along the driest expositions of the Southern slope of Keminskiy ridge, ridge Kok-Oirok, as well as Eastern end of ridge Kungei-Ala-Too, at the height of 1500-1800m above sea-level. Low-mountain biocenosis of steppe of the Northern type are vividly-expressed at the Southern slopes of Zailiyskiy Ala-Too ridge with different ecological conditions. In this biocenosis 20 species of ecotopes live. In biocenosis of low-mountain steppe of Chon-Keminskiy national natural park, in comparison to other species of mammals, frequently can be found tamarisk gerbil, and at night – eared hedgehog. Within agrobiocenosis of this biocenosis a wide spread of the following rodents is observed: house mouse, grey hamster, and mole lemming. During summer in populated areas common pipistrelle become active at evening-to-night periods, and their rapid flight can be observed. Mammals of this biocenosis are defined by certain bio-ecological characteristics, typical only for them
Mid-mountation biocenosis
of high-grass lawns
Mid-mountation biocenosis of high-grass lawns are surrounded by biocenosis of low-mountain steppe from below and their upped border adjoints to biocenosis of fir forests and fir bushes. These biocenosis are mostly expressed in mid-mountain areas at the slopes of ridge Kungei-Alatoo, however, sometimes they advance into slopes of ridges of Zailiyskoye Alatoo and Keminskiye mountains. In this biocenosis totally 29 species of ecotopes of mammals. The most frequently-found are Kyrgyz vole, forest mouse, Tjan-Shan shrew. In rocky-rubbly caves colonies of sharp-eared bats can be found. This narrow line of mid-mountain biocenosis of high-grass lawns, located between tow biocenosis, can also be called ecotone, since between borders of communities there is no natural border, and high density of mammals is observed there. 80 % of all mammal ecotopes of Chon-Kemin natural park live in mid-mountain biocenosis of high-grass lawns.
Biocenosis of fir forests and fir bushes
According to our observations, this biocenosis is spread in a broad line from East to West along the Northern slope of ridge Kungei-Alatoo, but in form of separate areas plantings of Tjan-Shan spruce can be found in depth of ravines of the Southern slope of ridge Zaaliyskoye Alatoo, specifically in ravine Karagailuubulak. Their lower border is located at height 1800 m above sea level, and to the East along with escalation of grounds the border is located at height 2000-2300 m above sea level, above which Tjan-Shan spruce can sometimes take shape of elfin wood. The main dominant in vegetation is Tjan-Shan spruce, pea tree. For this biocenosis 24 ecotopes of mammal species are typical, main of which being Tjan-Shan birch mouse, Tjan-Shan forest mouse that are endemics. In this biocenosis forest mouse, boar, and narrow-skulled vole are frequently found.
High-mountain biocenosis of forest-steppe and alpine wastelands. This biocenosis is expressed at the Northern slope of ridge Kungei-Alatoo at the height of 2900-3100 m above sea level to 2600-3300-3900 m above sea level. It is located in the Eastern part of National natural park Chon-kemin. From West to East ground escalation increases, and therefore areas of biocenosis in these regions are represented by firn fields and ice caps. In places of high concentration of winter snow and along edges of slops alpine different-grass lawns can be found. At heights from 3800 to 4700 m above sea level prevail rocks, rocky slopes, and mountain wastes, country rocks, morainic debris, snow and firn fields, and also permanent ice caps. In total 20 species of mammals can be found in this biocenosis. In comparison to other biocenosis, here narrow-skulled voles, silver voles, and grey marmots can be found more frequently.
Populations of large species of mammals migrate from one biocenosis to another in search for food due to unfavourable changes in weather conditions depending on season of year. They live in biotopes of each biocenosis, finding “space of life” of a species, and diverse special structure of these populations can be observed.
During expeditions, along with collection of animals for cariologic analysis, we have also carried out initial quantitative accounting of number of mammals. But, we should like to outline that the received data on population of mammals in each biocenosis does not reflect a complete number of their populations. Our observations should be considered as initial information on ecologically-dominant species of rodents, background species, and which species has a high efficiency. Bioecological specific features, genetics, behavior, and their dynamics, as well as biocenotic interaction between animals are not studied sufficiently.
Characteristic of cariotype of certain mammals that live in biocenosis
of National natural park Chon-Kemin
At current level of ecology development traditoinal approach proves insufficient, especially while studying stenotopic and eurytopic species of animals, as in this case cariotype can become if not the only, but most important indicator of diagnostics. Cariologic characteristics in combination with classic data on morphophisiological ecology provide for specification of stenotopic and eurytopic species within limits of geographic expansion. Thus, penetration of the results of comparative cariology into auto-ecology made it urgent to study inter-relation between tolerance and cytogenetic data of micropopulations of species. Therefore, we have studied cytogenetic features of certain mammals together with biocenosis of this park.
Common pipistrelle–Pipistrellus pipistrellus Schreber (Vespertilionidae, Chiroptera). The smallest of hand-winged, common pipistrelle is a synantrope animal, and it is widely spread in different populated areas that are defined by certain specific features of ecologic environment.
Data on cariotype of common pipstrelle is provided in works [8, 17], from which we know that diploid chromosome set of common pipstrelle of Polish population consists of 44 chromosomes, chromosome arm NF = 50. Autosomes of this cariotype consist of three groups of chromosomes: 3 pairs of metacentric, 1 pair of submetacentric, and 17 pairs of body-centric chromosomes.
We have studied cariotype of common pipistrelle of Chon-Kemin population that consists of 44 chromosomes. Autosome arms equal NFа = 50 (Fig. 1).

Fig. 1. Metaphase plaster and cariogramme of common pipistrelle (Pipistrellus pipistrellus Schreber) of Chon-Kemin population (2n = 44, NFа = 50)
Autosomes consist of 3 pairs of large metacentric, 1 pair of mid-size submetacentric and gradually-decreasing line of 17 pairs of acrocentric chromosomes. Gender chromosomes of male specimen consist of metacentric X-chromosome and submetacentric Y-chromosome. Comparing the received data with other bibliographic information, we can establish special-biotopicstability of cariotypes of common pipistrelle.
Grey hamster – Cricetulus migratorius Pallas (Cricetidae, Rodentia). Grey hamster is a widely-spread species, these animals can be found in ecologically-diverse biocenosis up to 3600 m above sea level. Its tolerance to biotope is eurytopic.
Cariotope of grey hamster was studied for the first time [18], and the data [19] shows that cariotype of Turkish population consists of diploid set of 2n = 22 chromosomes and includes groups of 6 pairs of meta- and submetacentric, 5 pairs (including large sub-body-centric X-chromosome) of sub-body-centric chromosomes. According to the data [9], cariotype of grey hamster of Jumgal population and Kegetin population has diploid set of chromosomes 2n = 22, chromosome arm NF = 44. Apart from modal number of chromosomes it revealed poly-diploid cells. These changes are related to tectonic nature of the mountain Tjan-Shan, where realms of the mentioned populations are located.
We have studied cariotype of grey hamster of Chon-Kemin population. Its cariotype consists of diploid set of chromosomes 2n = 22, chromosome arm NF = 44. Chromosomes in cariogramme form 3 groups. The first group has a gradually-decreasing line of metacentric chromosomes. The second group consists of 1 pair of submetacentric chromosomes, the third – of 5 pairs of sub-body-centric chromosomes. Gender chromosomes are not identified (Fig. 2).
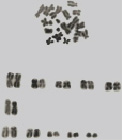
Fig. 2. Metaphase plaster and cariogramme of grey hamster (Cricetulus migratorius Pallas) of Chon-Kemin population (2n = 22, NF = 44)
While comparing the received results with bibliographic data we can establish stability of cariotypes according to morphology of chromosomes and underline relation between cariotype and ecological conditions of their habitat that allows us to use cariotype as bioindicator.
Regardless of high geographic and individually-physiological variability of Cricetulus migratorius, Pipistrellus pipistrellus, cariotypes of these species are stable in the most variable points of their realm. It seems that the main factor of high tolerance of certain mammals is stability of their cariotypes. Naturally, we can suppose that eurytopicity of species is determined by inheritance, and the studied species are widely-spread geographically due to high adaptive potential of their genotype, therefore, cariotypes of the studied populations are stable.
Thus, in modern researches on ecology of animal species data on comparative-cariologic analysis of inventory nature should be used widely in process of establishing eurytopic and stenotopic species.

Fig. 3. Metaphase plaster and cariogramme of Tjan-Shan mouse (Sicista tianschanica Salensky) of Chon-Kemin population (2n = 32, NF = 44)
Tjan-Shan birch mouse – Sicista tianschanica Salensky (Mammalia, Rodentia). A small rodent, it is an endemic species of mountain Tjan-Shan. This local species of mammals lives in hiding, and its biology is not widely studied. While analyzing cariotype of Tjan-Shan birch mouse of Chon-Kemin population we determined that diploid number of chromosome set, as well as that of Issykul population [11], equals 2n = 32. But, in comparison of chromosome morphology of both populations we discovered their morphological variability. Cariogramme was composed for each group along with gradually-decreasing line. Autosomes consist of 4 chromosome groups. The first group consists of 5 pairs of metacentric chromosomes, of them two large, two mid-sized, and the last pair – of small size. The next group consists of 4 pairs of submetacentric chromosomes, of them one large, others are mid-sized. The third group consists of 3 pairs of sub-body-centric chromosomes. Of them one large, one pair of mid-size, and the last is the smallest. The last group consists of 3 pairs of acrocentric chromosomes. Gender chromosomes consist of submetacentric and sub-body-centric. Chromosome set carries all types of morphologic chromosome structures. In each group chromosomes of different size can be found. We point out that according to the received data, cariotypes of isolated micropopulations of species Sicista tianschanica have spacial-biotopic chromosome polymorphism.
Tjan-Shan forest vole – Clethrionomys centralis Miller (Mammalia, Rodentia). It is a specifically mountain kind of voles. Tjan-Shan forest vole is an endemic of national natural park. According to our cariologic studies, cariotype of Chon-Kamin population of Tjan-Shan forest vole consists of diploid chromosome set 2n = 56. Chromosome arm NFa = 56. Morphology of gender chromosomes differs slightly from bibliographic data [10]: X-xhromosome is of sub-body-acrocentric size, Y-chromosome is same in morphology, but smaller in size. Comparison between cariotypes of Almatinskaya and Chon-Kemin population revealed chromosome polymorphism. The discovered smallest metacentric chromosomes of Almatinskaya population are not found in cariotype of Chon-Kemin population. Morphology of gender chromosomes also varies. However, diploid chromosome set is same and equals 2n = 56. According to the received data on cariotype of this mammal, we can outline that cariotypes of separate populations have spacially-biotopic chromosome polymorphism.

Fig. 4. Metaphase plaster and cariogramme of Tjan-Shan forest vole (Clethrionomys centralis Miller ) of Chon-Kemin population (2n = 56, NFа = 56)
Narrow-skulled vole – Microtus gregalis Pallas (Mammalia, Rodentia). This species lives in high-mountain areas of national natural park. Colonial animal. Cariotype of narrow-skulled vole of Chon-Kemin vole consists of diploid chromosome set 2n = 36. Autosome arm NFa = 50. Autosomes consist of 3 morphologic groups of chromosomes. The first group consists of 5 pairs of metacentric, the second group – 3 pairs of submetacentric, and the third group – 9 pairs of acrocentric chromosomes. All groups are composed along the decreasing line in their size. Geterochromosomes: X-chromosome is formed of large metacentric, Y-chromosome – of smaller acrocentric. In certain metaphase plasters in the 11 pair secondary constrictions were discovered. Cariotypes of narrow-skulled vole of Chon-Kemin population was compared to cariotype of (analyzed at department of general biology, ecology, and educational technologies of KSU of Z. Balasagyn, Kyrgyzstan) of Aksay population that consists of diploid number 2n-36, NFa = 50. The compared cariotypes of populations are different in morphology of autosomes, as well as in morphology of geterochromosomes. Thus, at the foundation of these comparisons we can observe special-biotopic chromosome polymorphism of cariotypes of narrow-skulled vole.

Fig. 5. Metaphase plaster and cariogramme of narrow-skulled vole (Microtus gregalis Pallas) of Chon-Kemin population (2n = 36, NFа = 50)
According to the carried out cariologic research on cytogenetics of certain mammals of Chon-Kemin population, we can generalize the data, presented above. Stability of cariotypes of common pipistrelle, grey hamster in number of chromosome set and morphology in all studied “areas of life” is observed. For Tjan-Shan birch mouse, Tjan-Shan forest vole, narrow-skulled vole variability of cariotypes is expressed in form of spacial-biotopic chromosome polymorphism.
Thus, in modern research on ecological cytogenetics of species data of comparative cytogenetic analysis of inventory nature should be used widely in establishment of eurytopic, stenotopic, and genetically-stable polymorphic species of animals.
Библиографическая ссылка
Sharshenaliyeva G.A., Beishenaliyeva S.T., Usengaziyeva G.S. BIOCENOSIS, BIOCENOTIC DIVISION OF MAMMALS IN NATIONAL NATURAL PARK CHON-KEMIN AND CHARACTERISTIC OF CERTAIN MAMMALS OF THIS PARK // European Journal of Natural History. 2019. № 3. ;URL: https://world-science.ru/en/article/view?id=33973 (дата обращения: 25.06.2026).